Apomixsis- definition, classification, Agamospermy, vegetative reproduction
APOMIXIS
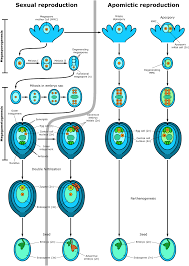
The normal sexual cycle (amphimixis) involves two important processes (a) Meiosis, this transforms a diploid sporophytic cell (megaspore mother cell, MMC) into four haploid gametophytic cells, and (b) Fertilization, In which two haploid gametes of opposite sex fuse re-establishing the diploid sporophytic generation. Thus, in a sexual cycle a diploid generation (sporophytic) alternates with a haploid generation (gametophytic) In angiosperms the gametophytic generations are very short and are represented by embryo sac on the female side and microspores or pollen grains on the male side. In some plants meiosis and syngamy are interrupted and still a viable embryo is formed within the confines of the seed-coat. Such asexual seeds, which produce progeny identical to the female parent, are called apomictic seeds, and the phenomenon is known as apomixis (Apo = away from + mixis = act of mixing or mingling). The initial discovery of apomixis in higher plants is attributed to the observation that a solitary female plant of Alchornea ilicifolia from Australia continued to form seeds in Kew Gardens in England (Smith, 1841). apomixis is more common in polyploids than diploids. Following Winkler (1908), who coined the term apomixis to mean "substitution of sexual reproduction by an asexual multiplication process without nucleus or cell fusion", plant breeders restrict the scope of apomixis to asexual seed formation (Bashaw, 1980; Khush et al., 1994). However, many authors favour a broader concept, according to which apomixis refers to substitution of the usual sexual reproduction by a form of asexual reproduction which does not involve meiosis and syngamy. Accordingly, there are two main categories of apomixis.

In this category the plants propagate by a part of their body other than the seed. The structural units employed for this purpose are called propagules.
2. Agamospermy:

The plants belonging to this category have retained seed as the agent of propagation but the embryo is formed by some process in which normal meiosis and syngamy have been eliminated. Two different types of agamospermy are recognized, based on whether the embryo develops from a cell of the unreduced female gametophyte (gametophytic) or directly from diploid sporophytic cells of the ovule, such an nucellus and integument sporophytic or adventive embryony In gametophytic apomixis an unreduced embryo sac develops from the MMC by circumvention of meiosis (diplospory) or directly from a cell be their nucellus (apospory). The embryo is formed by the unfertilized egg (parthenogenesis) or some other cell of the embryo sac (apogamety). Curtain species of Potentila show both apospory and apogamy In gametophytic apomixis alternation of gametophytic and sporophytic generations maintained.
(1) The propagules are formed outside the floral regions
and, despite the occurrence of functional sex organs, no fertilization or seed
setting takes place Agave americana and Elodea canadensis are examples of this type.
(2) The propagules are formed outside the floral regions and
the plants are sexually sterile. Fritillaria imperialis and Lilium bulbiferum
are typical representatives of this group. They propagate by means of bulbils
and bulblets. A species may have sexually sterile as well as sexually fertile
races (F. imperialis) The former shows stronger tendency for vegetative
propagation

AGAMOSPERMY
Adventive Embryony:
In this type of agamospermy the gametophytic generation is completely eliminated. In this respect it is close to the vegetative propagation, but differs from it essentially in two ways: (a) it has retained the seed habit, and (b) one or more diploid sporophytic cells undergo the stages of embryogeny and grow into mature embryos inside the normally developed sexual embryo sac. The zygotic embryo either degenerates or competes with the apomictic embryos.Adventive embryony usually leads to the formation of more than one embryo in a seed. Besides the well known examples of citrus and mango, adventive embryony is known to occur in Buxaceae. Cactacear. Euphorbiaceae, Myrtaceae and Orchidaceae
Gametophytic Apomixis:
Diplospory:
In this category of
apomicts, the MMC differentiates as in sexual ovules but it does not enter
meiosis. A special feature of apomictic megasporocytes is that they lack a
callose wall around them unlike the megasporocytes undergoing meiosis in sexual
individuals of the same species (Savidan, 2001). It produces diploid embryo sac
through mitotic divisions As early as 1896, Juel described diplospory in
Antennaria al Since then this phenomenon has been reported in several species.
Of the plants known to exhibit gametophytic apomixis, 75% belong to three
families, the Asteraceae, Rosaceae and Poaceae (Bicknell and Koltunow, 2004).
Four types of diplosporic embryo sac development have been described, and named
after the genera in which they were first observed
The MMC enters
meiotic prophase and pairing of homologous chromosomes occurs to some extent.
However, due to precocious desynapsis univalents are scattered over the spindle
at metaphase-1 A restitution nucleus is formed after the first meiotic
division. The MMC with the restitution nucleus divides mitotically to form a
dyad with somatic (2n) chromosome number Usually the micropylar cell of the
dyad degenerates while the chalazal cell undergoes further mitotic divisions to
form an 8-mucleate embryo sac. Besides Taraxacum, type of diplospory occurs in Erigeron
sp., Aralus halbellis and Agropyron scabra
(ii) Ixeris type:
In the triploid races of Ixeris dentata the MMC divides by a
semi-heterotypic (asyndetic) meiotic prophase, in which there is no pairing and
a restitution nucleus results. The restitution nucleus undergoes 3 mitotic
divisions leading to the formation of an 8-nucleate embryo sac.
(iii) Antennaria type:
MMC does not enter
meiosis and acts as an unreduced megaspore After a long interphase, it
increases in size and shows pronounced vacuolation to appear like functional
megaspore Through mitotic divisions it forms an unreduced 8 nucleate embryo sac
of Polygonum type This type of diplospory is of wide taxonomic distribution. It
has been reported in Tripsucum dactyloides, T. zopilotense
and Eupatorium glandulosum (Leblanc and Savidan, 1994).
(iv) Allium Type:
In this type of premeiotic chromosome doubling is the cause of unreduced embryo sac formation (Hakansson and Levan 1957; Kojima and Nagato, 1992). Chromosome number in MMC is doubled by endomitosis and the ensuing meiosis results in a dyad of unreduced cells. Two subsequent mitoses in the chalazal dyad cell results in an 8-nucleate embryo. sac Allium nutans and A. odorum show Allium type of diplospory. Kojima and Nagato (1992) reported 76%-98% diplospory in six cultivars of A. tuberosum, based on the percentage of reduplicated MMC.

Apospory:
Apospory is by far
the most common mechanism of apomixis in higher plants (Bashaw, 1980). It was
first reported by Rosenberg (1907) in Hieracium sp. Within the genus Hieracium,
subgenus Hieracium. H murorum is autonomous diplosporous apomict whereas the
subgenus Pilosella species H pracaltum exhibits autonomous apospory, Obligate
ucellar apospory with pseudogamy has been observed in seven species of Pennisetum
P macerum meum , Poritair P pedicellation plantace, and Pamulatum (Dujardim and
Hanna, 1 199) In aposporous plants a normal MMC differentiates in the sa manner
as in sexual ovules and begins to enter meiosis which may or m not complete
both divisions to form a linear tetrad. At some stage during this period one or more
adjacent nucellar cells become meristematic appear like MMC These cells undergo
mitotic divisions to form unreduced mitotic embryo sacs. In obligate apospory
the sexual MMC or its products ab and the sporous embryo sac occupies the
entire area of the ovule The aposporous of degeneration timing of initiation of
the sexual embryo sac depends on the time of aposporous embryo sacs The sooner
an aposporous initial induced earlier the degeneration of the developing
megaspore occurs. There in considerable variation, even within the same
species, with respect to de number of fully developed aposporous embryo sacs
formed within an ovule. In the aposporous members of the Asteraceae, only one
nucellar cell acts the mother cell and gives rise to structurally normal,
8-nucleate embryo sac In grasses, on the other hand, more than one embryo sac
may develop in the same nucellus with 4-nucleate monopolar organization. In
aposporous H. aurantiacum of the subgenus Pilasella, multiple embryo sacs in an
ovule fuse into one due to degeneration of their walls during their development
(Koltunin et al, 1998, 2000) Occasionally, aposporous and sexual embryo sacs
may co-exist Apospory is mainly of two types
In Hieracium, subg Pilosella, the MMC undergoes the usual meiotic division and forms a tetrad. At this stage a nucellar cell at the chalazal end of the tetrad becomes activated and starts developing into an aposporous, unreduced embryo sac All the four megaspores gradually degenerate and the aposporic embryo sac matures. Occasionally, aposporous embryo sac may develop in addition to the reduced embryo sac. This type of aposporic embryo sacs are 8-nucleate.

(i) Panicum type:
In Panicum and some other grasses, the aposporous embryo sac is 4-nucleate, organized into a 3-celled egg apparatus and a polar nucleus Antipodals are absent In Poa pratensis the ovules contain multiple embryo sacs but only one of these gives rise to an embryo in the mature seed (Savidan, 2001). Generally the largest embryo sac in an ovule, which is closest to the micropyle, is fertilized and forms seed.
REFERENCE :
THE EMBROLOGY OF ANGISPERMS 6th EDITION Author; SS BHOJWANI, SP BHATNAGAR ,PK DANTU.


Comments